Introduction | Discussion of Data | Elements | Discussion of Classification | Research Program | References | Feedback
The Malva alliance is a group of related genera forming part of subtribe Malvinae, tribe Malveae and subfamily Malvoideae, of the family Malvaceae sensu APG. The latest revision of Malvaceae, by Kubitzki & Bayer [3], includes 4 genera - Malva, Lavatera, Alcea and Althaea - in the alliance, and places Malope and Kitaibelia in a separate Malope alliance. Elsewhere separate Malva and Althaea alliances have been proposed.
These genera are the only elements of subtribe Malvinae native to the Palaeoarctic. However they also extend to the Ethiopian (Lavatera abyssinica in the Horn of Africa, and Althaea ludwigii in southern Africa), Oriental (Malva sylvestris in China and Himalayan India, and Malva verticillata in China), Australian (Lavatera plebeia in the southern half of Australia, and Althaea australis known from a single collection in South Australia) and Nearctic (4 species of Lavatera on Californian and Baja Californian islands) regions, and several species are widely naturalised in temperate and tropical highland regions throughout the world.
DNA sequences deposited in GenBank for the Internally Transcribed Spacer (ITS) and 5.8s nuclear ribosomal RNA (5.8s) loci place Althaea (section Althaeastrum) and Alcea nearer to Kitaibelia than to Malva and Lavatera. Unpublished ITS data from Escobar et al [5] places Malope trifida nearer to Malva and Lavatera than to Althaea (section Althaeastrum) and Alcea (contrary to the expectation from morphology and biogeography that Malope is the sister genus to Kitaibelia). Tate et al [12] sample both Malope (Malope malacoides) and Kitaibelia, and confirm that these two genera do not form a monophyletic group.
Thus Kubitzki & Bayer's concept of the Malva alliance is paraphyletic with respect to their Malope alliance. Here I divide the 6 genera into 2 alliances; a Malva alliance composed of Malope, Malva and Lavatera, and an Althaea alliance composed of Althaea, Alcea and Kitaibelia. I have considered DNA evidence to indicate that the 6 genera form a clade - the Eurasian Malvinae - to which the sister group might be the South African genus Anisodontea, giving an Old World Malvinae clade; however the results of Tate et al [12] have Anisodontea and the North American Callirhoe alliance (Callirhoe and Napaea) nested within this clade.
The Malva alliance as here constituted was traditionally divided into the three genera Malope, Malva and Lavatera, the first consisting of those species with capitate fruits and cordate bracteoles, the second of those with unfused bracteoles, and the last of those with fused bracteoles. There is evidence that this latter division is untenable. Firstly there are single species (L. mauritanica and L. triloba) in which fusion of bracteoles differs between populations; secondly it is inconsistent with DNA sequence data deposited in GenBank; and thirdly it is inconsistent with the distribution of other character traits.
The interpretation of ×Malvalthaea transcaucasica as a hybrid between Malva aegyptia and Althaea hirsuta suggests that these species and their allies are relatively closely related; either one group being misplaced in Malva or the other in Althaea. Krebs [8] proposed that Althaea section Hirsutae was properly placed in the Malva+Lavatera clade (he proposed a segregate genus Dinacrusa, based on Alefeld's subgenus of that name, with contents as given for section Dinacrusa below). The ITS data of Escobar et al [5] confirm this location, but with Krebs' Dinacrusa paraphyletic with respect to the remainder of Malva and Lavatera (excluding section Navaea).
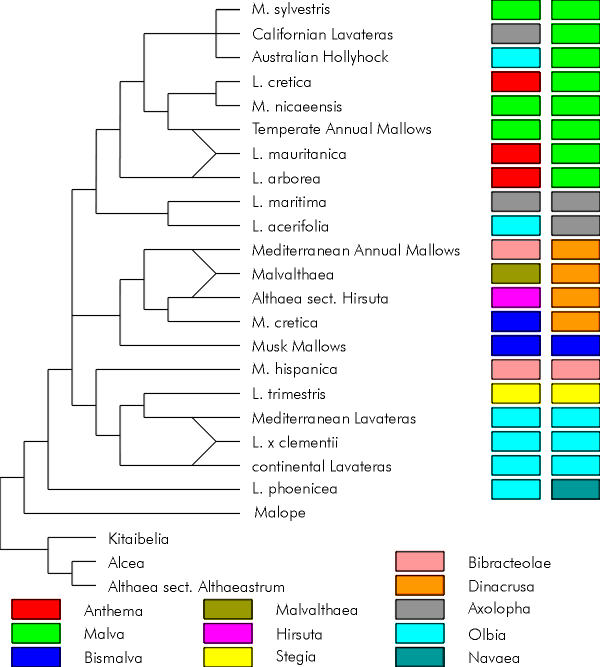
The following tree is based on the cladograms of Ray [9, 10], Escobar et al [5] and Tate et al [12], and is provided as a basis for discussion below. The two columns show "traditional" and proposed divisions of the alliance into sections.
The above tree is a combination of those presented by Ray [9, 10], Escobar et al [5] and Tate et al [12]. Not all taxa are explicitly placed on the cladogram.
Traditionally Malva and Lavatera were distinguished by epicalyx characters: Malva has 3 (or 2) unfused bracteoles, and Lavatera 3 fused bracteoles. (Some old literature describes the epicalyx of Lavatera as being 3–6-fid. This may be because the shape of the 3 fused bracteoles in some species results in a 6-pointed epicalyx.) This division was long recognised to be problematic [a, 5, 6], and Ray [9, 10] found it to be untenable. I haven't seen Ray's dissertation [b], but his principal distinction seems to be between species with malvoid mericarps (i.e. dorsal faces flat) and those with lavateroid mericarps (i.e. dorsal faces rounded). This distinction fails for M. tournefortiana (Bismalva), which may have either rounded or flat dorsal faces.
Althaea section Hirsutae has 9 fused bracteoles, alternatively interpreted as 3 fused bracteoles, each 3-lobed. ×Malvalthaea has 3 or more bracteoles, which on the preceding interpretation would be 3 fused bracteoles, variously lobed.
Wiggins [1] states that Lavatera has articulated pedicels, and Malva unarticulated pedicels. This would correlate with unfertilised flowers abscissing at the articulation in the former, and the flower and pedicel shrivelling and falling as a unit in the latter. The state of this trait is not known to me for many taxa, but M. moschata has an articulated pedicel, though flowers do not always absciss there, and L. cretica and L. mauritanica appear to lack an articulation. However, I have observed an articulation of the pedicel in the sterile (suspected hybrid) cultivar Malva sylvestris 'Marina'.
The ITS DNA data deposited in GenBank support the unity of the Malva alliance. With the exception of Malope, they share two considerable deletions in ITS and some point substitutions in the relatively conserved 5.8s nrRNA sequence. Section Navaea is conservative, and section Malva derived; other sections for which information is available are intermediate.
Leaves moderately heterophyllous, lower leaves orbicular, upper leaves 3–5-lobed, flowers borne singly in leaf axils, held ascending, long-pedicelled; bracteoles large and cordate; staminal column sometimes bearing 5 distinct multiranked columns of filaments; schizocarp capitate.
Leaves palmately lobed, degree of heterophylly not known to me; flowers borne in pedunculate clusters [2, 4] of 3 to 5 in the leaf axils, epicalyx caducous; mericarps elongated reniform, with 2 apical spines [7], more numerous (ca. 30-40) than in other sections of Malva and Lavatera.
A single rare species from the Canary Islands. In the DNA dataset it is conservative and relatively isolated, being the sister group to the remainder of the alliance. Details of the species are hard to come by, but although de Candolle [4] and Baker [2] both place it in section Olbia they give descriptions which include the distinctive characteristics given above.
Foliage strongly heterophyllous (except L. oblongifolia and L. triloba), lower leaves cordate-orbicular, variously lobed, upper leaves more distinctly lobed, then hastate and finally lanceolate; inflorescence an initially corymbose leafy to bracteate raceme, flowers borne singly (except in L. triloba) in leaf axils, held ascending, short-pedicelled; fruit lavateroid.
The DNA sequences separate the continental species, Ll. cashmeriana and thuringiaca, from the Mediterranean species, Ll. olbia, punctata and triloba, the latter being closer to L. trimestris; DNA sequences aren't available for the remaining species. Morphologically L. olbia etc are closer to L. thuringiaca than to L. trimestris. Both this, and the existence of the hybrid L. ×clementii (olbia × thuringiaca) suggest that the DNA sequences are misleading in this case, and I retain these species in a single section. It is possible that the discordance is the result of ancient hybridisation.
Lavatera triloba is aberrant in a number of characteristics, but is placed near to L. olbia by DNA sequence data. L. thuringiaca often displays a cuspidate epicalyx similar to that of L. trimestris.
Foliage moderately to strongly heterophyllous, lower leaves cordate-orbicular, 5-angled, upper leaves more distinctly lobed, finally cuneate-hastate; stems with deflexed hairs; inflorescence an initially corymbose leafy to bracteate raceme, flowers borne singly in leaf axils, held ascending, long-pedicelled; epicalyx cuspidate; fruit lavateroid; fruit axis expanded to cover the mericarps.
1 species, the annual Lavatera trimestris, which may be distinguished by the swelling of the fruit axis to cover the mericarps, and by several other traits.
ITS data does not clearly separate Lavatera trimestris from section Olbia, Lavatera punctata (including Lavatera maroccana) particularly associating with Lavatera trimestris. However the phylogeny here is probably complicated by reticulatory hybridisation, and I conservatively retain Stegia as a separate section.
Foliage moderating heterophyllous, lower leaves orbicular, upper leaves sub-rhomboid, bracteoles short (c. 3mm).
Malva hispanica has traditionally been placed in sect. Bibracteolatae (sect. Bismalva s.l), with Malva aegyptia and Malva trifida, on the basis of its possession of only two bracteoles in the epicalyx. It doesn't agree with those species in other traits, other than its annual habit. The pattern of heterophylly bears similarities to that of Lavatera section Olbia, the lower leaves being orbicular, and the upper leaves sub-rhomboid, with which it also share a numbers of mericarp traits.
The ITS data of Escobar et al [5] place it as sister to a clade consisting of sections Olbia, Stegia, Axolopha and Malva. I tentatively assign it to a monotypic section; as Malva hispanica is the first taxon in De Candolle's circumscription of section Bibracteolae [4], I believe that the sectional name follows this species, rather than the other bibracteolate mallows, Malva aegyptia and Malva trifida, and therefore Bibracteolae is the appropriate name for the section. Whether Malva or Lavatera is the best home for this is not clear.
Foliage variably but usually strongly heterophyllous, lower leaves orbicular, scarcely lobed, upper leaves often digitate with pinnatifid lobes, sometimes lobed and strongly toothed (e.g. Malva alcea 'Fastigiata'); inflorescence an initially corymbose raceme, lowest flowers in leaf axils, the remainder subtended by short stipule-like bracts; petal veins not constrasting in colour with remainder of petal; petal apices erose; fruit lavateroid or malvoid (some individuals of M. tournefortiana); perennials.
The musk mallows, MM. alcea, moschata and tournefortiana are an obvious close grouping. MM. alcea and moschata are easy to hybridise, though the hybrid (M. ×intermedia), being triploid, is nearly completely sterile.
Foliage heterophyllous, similar to section Bismalva; epicalyx of 2 free bracteoles, mericarps
Foliage heterophyllous, similar to section Bismalva; epicalyx either of 2 free bracteoles, 3 free bracteoles or of 9 fused bracteoles; mericarps lavateroid, malvoid (Malva cretica) or keeled on dorsal face; annuals.
This is a difficult group, consisting of species previously placed in Malva and Althaea. The existence of the hybrid × Malvalthaea transcaucasica (A. hirsuta × M. aegyptia) implies that one of the species, and by implication its allies, is misplaced. However, either would be aberrant in the other genus. Furthermore, given the reported existence of relatively wide hybrids within the alliance, such as M. × inodora (M. moschata × sylvestris) and M. ×egarensis (M. alcea × sylvestris), it cannot be assumed that the group is monophyletic.
The ITS tree of Escobar et al [5] places Malva cretica as sister to the annual Althaeas. Otherwise this tree has these species as a paraphyletic assemblage with respect to sections Bismalva, Olbia, Stegia, Axolopha and Malva. This is apparently inconsistent with the existence of ×Malvalthaea transcaucasica, which on their tree would be nearly the widest possible hybrid in the Malva alliance. (I am not familiar with the evidence for the hybrid nature of ×Malvalthaea transcaucasica.)
Foliage orbicular and lobed, scarcely heterophyllous; flowers held slightly nodding, borne solitarily, or less commonly in pairs, in leaf axils, not in a identifiable raceme; malvoid fruit; shrubs or subshrubs.
Two species, not particularly close, but which are sisters within the DNA sequence data. There are morphological characteristics to support the sister group relationship.
Foliage orbicular (distinctly lobed in Malva arborea, Malva assurgentiflora and perhaps others), moderately heterophyllous, upper leaves more distinctly lobed; inflorescence an initially corymbose leafy raceme; flowers fasciculate in leaf axils (solitary in Malva occidentalis), held upright, petals narrow, few veined, veins often prominently coloured; mericarps malvoid, i.e. dorsal face flat. Habit varied, prostrate or erect annuals, biennials and herbaceous perennials, shrubs and small trees.
Although the most diverse part of the Malva alliance, this group is also the best supported section. In the ITS data it can be divided into 3 subgroups, Malva arborea, with a strongly fused epicalyx, and distinctive subarborescent habit, the perennial species (MM. sylvestris, assurgentiflora, venosa, occidentalis and lindsayi) with Lavatera abyssinica and the biennial Malva australiana, and the annual species (MM. neglecta, nicaensis, parviflora, pseudolavatera, pusilla, vidali and verticillata). There is insufficient data to place Malva microphylla, Malva bucharica and Malva pamiroalaica in particular subgroups. Malva durieui appears to be of polyploid hybrid origin between Malva arborea and one of the annual species; cpDNA data from Escobar et al [5] suggests that Malva parviflora was the seed parent.
This group is supported by cross-breeding experiments, and field observations of hybridisation, in addition to DNA sequence and morphological data. Good seed set is often obtained with crosses between species in this group, while if seed is obtained with crosses with sections Olbia and Bismalva seed set is always poor. In an experirmental hybridisation program Kristofferson [15] found that crosses between Malva verticillata ("section Conocentrae") and the remaining species traditionally placed in Malva ("section Planocentrae") were highly sterile, while crosses within the latter group, while having reduced fertility, have appreciable seed set. This suggests a different division to that found in the ITS data.
Malva leonardii, as described in Flora Iranica [11], is similar to section Malva in fruit characters, but has more deeply lobed leaves, and pedunculate, rather than fasciculate, axillary inflorescences.
It would appear that the Malva alliance sensu strictu should either be united into two genera (i.e. Malva and Malope), or divided into 4 or more genera.
The advantage of a two genus classification is that Malva would then represent a single monophyletic group. Furthermore Malveae is perhaps already oversplit, compared to Hibisceae. The disadvantage is that there are no clear characteristics which distinguish that group as a whole.
The advantage of a multiple genus classification is that morphologically well defined monophyletic groups can be defined. There are several problems; firstly which sections are joined together to make genera; secondly the position of section Dinacrusa and some species is unclear; and thirdly the major morphological breaks do not align with the phylogenetic breaks as indicated by ITS DNA sequences.
It seems clear that section Navaea (L. phoenicea) can be separated off, as originally proposed by Webb [d], and it would seem sensible to combine sections Olbia and Stegia in a genus Lavatera. Section Bismalva agrees with Lavatera morphologically (strongly heterophyllous, non-fasciculate inflorescences, lavateroid mericarps) more than with section Malva (unfused bracteoles), but the ITS sequences are ambiguous; depending on the choice of outgroup it is either sister to Malva, or to the combination of Malva, Olbia and Stegia. On the current data, to retain monophyly Medik's genus Bismalva would have to be resurrected, with sections Axolopha and Malva being retained in genus Malva. The position of "section" Dinacrusa remains unclear; I'm tempted to follow Fernandes [6] in uniting it with Bismalva, as in the tree shown above, but the evidence is insufficient. The appropriate location for Malva hispanica is also unclear.
Given the difficulties, unless a two genus classification is to be adopted, it would seem to be premature to formally reclassify the Malva alliance sensu strictu. As always, more research is needed.
Research on the correct classification of the Malva alliance can take 3 forms.
1. Floras and monographs generally do not describe sufficient traits for a useful cladistic analysis. I am building a set of detailed technical descriptions of specimens of Malvaceae as part of my Digital Herbarium, but this is unlikely to have a sufficiently wide coverage of taxa to be useful in the foreseeable future. Please contact me if you can collaborate with me on the Digital Herbarium, or can provide propagation material.
2. I am hybridising species within the Malva alliance, subject to constraints of availability and flowering season. Again I am limited in my coverage of taxa. Please contact me if you have any observations of hybridisation (including failures), or can provide propagation material. (I elsewhere describe known hybrids in the Malva alliance.)
3. The current available data set consists of ITS and 5.8s RNA sequences, mostly produced by M.F. Ray. The data set can be improved in two ways; firstly by adding sequences for additional loci; and secondly by adding sequences for additional taxa (unsampled taxa are Lavatera stenopetala, Lavatera microphylla, Lavatera vidali, Malva pusilla and Malva verticillata; sampling of geographic variants of the other species could also be performed). Use of the loci used by Jennifer Tate in her study of Tarasa etc [14] would be synergistic, in that it would also shed light on the relationships between the various alliances in Malvinae. Alternatively a suitable nuclear DNA locus might shed light on reticulation.
I am not in a position to do the sequencing myself (in the current state of sequencing technology it might be suitable for an undergraduate project), but could provide plant material, subject to the restrictions of phytosanitary regulations.
Escobar et al [5] have also sampled cpDNA sequences (psbA-trnH and trnF-trnL loci). The results are not concordant with the ITS sequences. An RFLP analysis of cpDNA by Fuertes et al [13] is also discordant with the ITS data, but in a different way. The former has Althaea section Althaeastrum and Alcea nested within the Malva alliance as here defined. The latter has Malva canariensis sister to Malva assurgentiflora, rather than Malva wigandii, and Malva wigandii nearer to the base of the tree, rather than as sister (with Malva canariensis) to section Malva.
If you have found any errors on this page, or can provide any further information then please contact me at botany@malvaceae.info.
© 2003, 2004, 2005, 2007, 2008, 2009 Stewart Robert Hinsley